

Bases
Morfológicas e Funcionais do Músculo esquelético
Autores:
Dionisio
V.C.
Descrição:
Este
texto trata dos principais aspectos morfológicas e funcionais do
músculo esquelético para a prática fisioterapêutica.
Temas:
Estrutura
e função muscular
Palavras-chave:
Músculo
A
fisioterapia tem como uma das suas principais ações, o trabalho de
treinar o músculo esquelético, seja produzindo alongamento, seja
para aumentar a sua capacidade funcional. Portanto, a compreensão
dos principais aspectos morfológicos e funcionais do músculo
esquelético é importante para que o fisioterapêuta possa eleger e
aplicar as estratégias de treinamento no músculo esquelético. Este
texto envolve os aspectos mais importantes das bases morfológicas e
funcionais do músculo esquelético e o maior aprofundamento do tema
exige a leitura de outros materiais, incluindo as referências que
serviram de base para este texto.
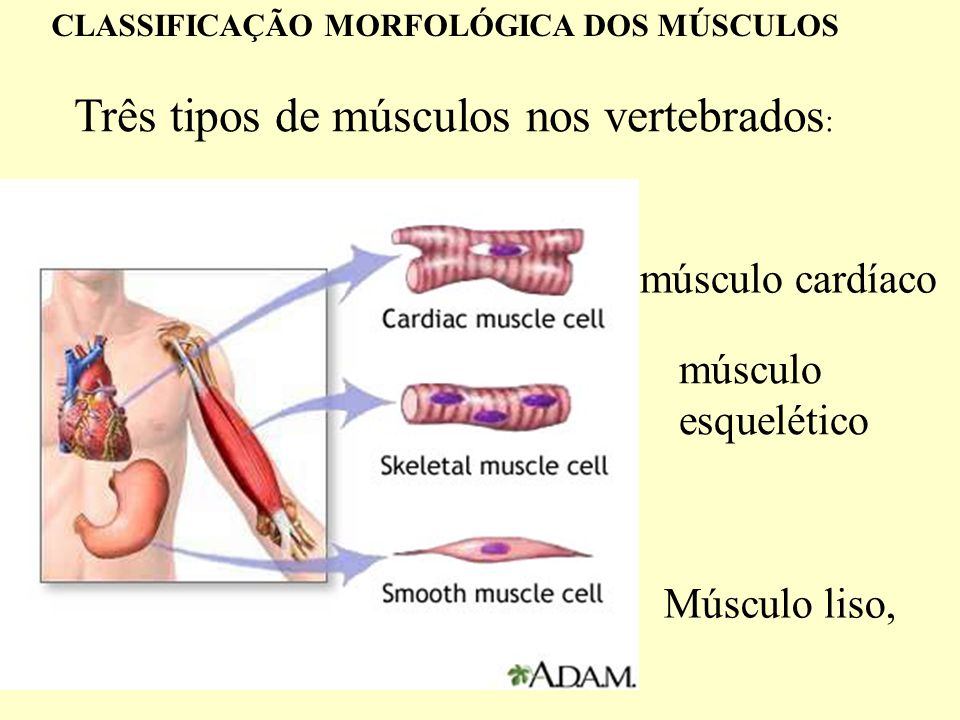
Os músculos e a gravidade são os principais produtores do movimento humano. Os músculos são usados para manter uma posição, levantar ou abaixar uma parte do corpo, desacelerar movimentos e para gerar velocidade no corpo ou em um objeto. Os músculos também são responsáveis por gerar forças compressivas nas articulações aumentando sua estabilidade. Os músculos são arranjados de forma que possam contribuir individualmente ou coletivamente para produzir um movimento muito pequeno ou um muito amplo e potente. Eles movimentam os ossos nas articulações e dão sustentação à postura do corpo contra forças. Portanto, é um dos principais objetos de trabalho do fisioterapeuta, sendo essencial a compreensão de sua base estrutural e funcional.
O músculo esquelético é o mais abundante no corpo humano com cerca de 40% do peso do corpo. A unidade estrutural do músculo esquelético é a fibra muscular, que consiste de muitas miofibrilas revestidas de uma membrana chamada de sarcolema. O sarcolema é conectado com as linhas Z por proteínas ricas em vinculina e distrofina.
A miofibrila é composta de vários sarcômeros que contêm filamentos de actina, miosina, titina e nebulina. Cada fibra muscular é envolvida por um tecido conjuntivo chamado de endomísio. As fibras musculares são organizadas em feixes, chamados fascículos, que são envolvidos por uma bainha de tecido conjuntivo conhecida como perimísio. O músculo é composto por vários fascículos envolvidos por uma fáscia de tecido conjuntivo fibroso chamado epimísio.
Geralmente, a extremidade de cada músculo é conectada aos ossos por tendões, os quais não possuem propriedade contrátil ativa. Os tendões são contínuos as fibras de colágeno do perimísio e epimísio. Os músculos formam o componente contrátil e os tendões os componentes elásticos em série. Já o endomísio, perimísio, epimísio e o sarcolema compõem os componentes elásticos em paralelo.
Quando o músculo é estirado passivamente, ocorre aumento de tensão e há armazenamento de energia pelos elementos elásticos em série e em paralelo. Por outro lado, quando cessa a tensão a energia é liberada. A possibilidade de distender e a elasticidade dos componentes elásticos são importantes para assegurar a transmissão suave da tensão durante a contração; retorno à posição original após a contração; prevenção de sobre estresse nos elementos contráteis; e a absorção e dissipação de energia.
Cada fibra muscular é composta de muitas miofibrilas acomodadas em paralelo umas as outras no sarcoplasma. Um padrão estriado se repete nas miofibrilas e que é referente aos sarcômeros. O sarcômero é a unidade funcional do sistema contrátil. Ele é composto de filamentos finos (composto de actina); filamentos grossos (composto de miosina); filamentos elásticos (compostos de titina); e componentes inelásticos (compostos de nebulina e titina).
A actina tem a forma de uma hélice dupla (fileira de gotas) na banda I. Duas proteínas adicionais estão junto à actina. A Tropomiosina é uma cadeia longa de polipeptídios que se acomoda nos sulcos entre as hélices da actina. A Troponina é uma molécula globular anexa periodicamente a tropomiosina.
A actina é conectada da linha Z (elemento que une os filamentos finos de sarcômeros adjacentes) até o centro do sarcômero sobrepondo-se aos filamentos grossos.
A miosina (filamento grosso) fica situada na região central do sarcômero, onde seu arranjo paralelo dá origem à banda escura (banda A). Ela é composta de moléculas com cauda e cabeça. Elas estão dispostas em direções diferentes a partir de cada metade do filamento. No centro do filamento há um espaço livre de cabeças (zona H). A titina liga a miosina à linha Z, se estendendo até a linha M (no centro da zona H, onde há união dos filamentos grossos), mantendo a posição central desse filamento. Da linha Z até a miosina a titina tem a aparência de uma mola, útil no desenvolvimento da força muscular passiva no estiramento. Ela também pode servir como um molde para a formação de miosina.
Os filamentos de nebulina se estendem da linha Z até os filamentos de actina, e podem servir como um molde para a formação de actina.
Junto às miofibrilas encontra-se acomodada uma rede organizada de túbulos e bolsas conhecida como retículo sarcoplasmático. Essa rede tende a alargar-se e fundir-se no nível das bandas A e I, formando as cisternas terminais e o sistema T (sistema transverso tubular).
A teoria proposta por Huxley parte do pressuposto que o sarcômero encurta por meio do deslizamento dos filamentos grossos e finos mantendo seu comprimento original. O deslizamento ocorreria por meio de pontes cruzadas (ligações entre actina e miosina) com posterior movimento angular da cabeça de miosina (similar ao remo), que depois se separa e se junta a outro local para novo movimento angular da cabeça.
As ligações cruzadas são assincrônicas, com cerca de 50% das ligações gerando força, enquanto os outros 50% manterão a posição quando os primeiros 50% se separarem.
O Cálcio é o mecanismo chave para a contração muscular. Se disponível, ocorre contração, mas se ausente a contração cessa. A presença do cálcio é regulada por eventos elétricos no sarcolema. O neurônio motor estimula o músculo (ao nível da junção neuromuscular) e o potencial de ação propaga-se ao longo de todo o sistema T. Um neurônio motor e as fibras musculares inervadas por ele formam a unidade motora, e respondem a lei do tudo ou nada (despolariza ou não despolariza). A quantidade de fibras para cada neurônio em um músculo depende de sua função, quanto maior a necessidade de precisão da ação muscular menor a quantidade de fibras musculares para cada neurônio. Os terminais axônicos acomodam-se nas fendas do sarcolema. Esses terminais estão repletos de vesículas sinápticas, as quais liberam a acetilcolina para transmitir o potencial de ação gerado no nervo para o sarcolema, e por fim, a liberação do cálcio.
A resposta mecânica ao estímulo nervoso é chamada de tetania. Existe um período entre o estímulo e a tetania, chamado de latência. Essa latência ocorre devido ao tempo necessário para eliminar a “folga” dos elementos elásticos. Ele também é chamado de atraso neuromecânico (também chamado de eletromecânico, no inglês electromechanical delay).
Do início da tetania até o seu pico temos o tempo de contração, e do pico até o final da tetania temos o tempo de relaxamento.
O acréscimo de resposta mecânica a estímulos sucessivos é chamado de somação. Se houve um segundo estímulo antes do término da tetania, haverá nova tensão, e possivelmente maior do que a primeira. Quando o segundo estímulo é logo após o primeiro (pequeno intervalo), há aumento da tensão resultante é maior do que seria com apenas um estímulo. Caso haja uma seqüência de estímulos com pequeno intervalo, ou seja, a freqüência é alta, haverá uma contração tetânica máxima, mantendo a tensão de pico.
As diferenças na gradação, freqüência de estímulo e número de unidades ativadas produzem os movimentos suaves que costumamos observar. A contração muscular exerce uma força sobre os ossos conhecida como tensão muscular. A força externa aplicada no músculo é chamada de resistência ou carga. A tensão muscular gera um efeito de rotação nas articulações chamado de momento ou torque. Ele é calculado como o produto da força muscular e a distância perpendicular da aplicação da força e o centro do movimento.
Contrações musculares e o trabalho muscular podem ser classificados de acordo com a tensão muscular e a resistência a ser superada. O trabalho estático ocorre quando nenhum trabalho mecânico é executado, mas há trabalho muscular (gasto de energia) por meio de uma contração isométrica.
O trabalho dinâmico ocorre quando um trabalho mecânico é produzido, e o movimento articular é gerado por uma das formas de contração muscular. A contração concêntrica ocorre quando os músculos geram tensão suficiente para superar a resistência do segmento do corpo e os músculos se encurtam. A contração excêntrica ocorre quando os músculos não geram tensão suficiente e são superados pela carga externa, de modo que eles se alongam (afastam origem e inserção). A contração isocinética é uma forma de trabalho muscular onde a velocidade de alongamento ou encurtamento se mantém constantes. Este tipo de contração é obtido por meio de equipamentos computadorizados. A contração isotônica é usada para definir a contração muscular com a tensão constante ao longo da amplitude articular. O uso desse termo não leva em conta que durante a amplitude de movimento ocorre mudanças no braço da alavanca, modificando também a tensão. Portanto, uma contração isotônica no verdadeiro sentido não existe, mas o termo muitas vezes é usado para definir uma contração que provoca movimento articular.
A tensão varia com o tipo de contração. As contrações isométricas produzem maior tensão do que contrações concêntricas. Contrações excêntricas podem gerar maior tensão do que contrações isométricas. Isto é aceito levando em conta a tensão suplementar produzida pelos elementos elásticos.
A classificação funcional da atividade muscular é necessária para melhor compreender como os músculos estão atuando. Agonista é o termo usado para denominar o músculo ou grupo muscular responsável por determinado movimento. O grupo muscular ou músculo responsável por realizar o movimento oposto ao agonista é chamado de antagonista. Quando um músculo auxilia o agonista na execução de um movimento é chamado de sinergista. Os fixadores são músculos que atuam com certo grau de sinergismo, uma vez que eles são músculos que produzem estabilidade em uma articulação de forma a favorecer o movimento em outra articulação.
Dependendo da posição e direção da resistência a superar ocorre diferenças na atividade do músculo, se concêntrica, excêntrica ou ainda não participa da atividade.
A produção de força ou tensão de um músculo depende do comprimento em que o mesmo está quando estimulado. A tensão máxima é produzida quando o músculo está próximo do seu comprimento de repouso. Quando o músculo está em uma posição encurtada, a tensão cai lentamente a princípio e depois mais rapidamente. Quando o músculo está em uma posição de alongamento, a tensão progressivamente diminui. Isto está relacionado com as alterações estruturais no sarcômero. Na posição de repouso, os filamentos de actina e miosina estão sobrepostos ao longo de todo o seu comprimento e o número de pontes cruzadas é máximo. Na posição alongada, haverá menos haverá menos pontes cruzadas e a tensão será menor. Na posição encurtada os filamentos estão sobrepostos nos terminais opostos do sarcômero, diminuindo a tensão.
A velocidade de encurtamento é maior quando a carga externa zero, mas à medida que a carga aumenta, a velocidade diminui. Quando a carga é aumentada ainda mais, o músculo alonga-se e contrai excentricamente. Com o aumento da carga ocorre maior velocidade de alongamento. Portanto, a relação carga-velocidade da contração excêntrica é inversa à da contração concêntrica.
A força ou tensão é proporcional ao tempo de contração. A força é maior se há tempo necessário para a tensão produzida ser transmitida para os elementos elásticos. O músculo que apresenta fibras musculares com muitos sarcômeros em série, terá maior velocidade e excursão (alcance de trabalho). O músculo que apresenta maior número de sarcômeros em paralelo terá maior produção de força.
O músculo realiza maior trabalho se for alongado e logo em seguida realizar uma contração concêntrica, a partir de uma contração isométrica. Os exercícios pliométricos são um bom exemplo desse efeito do pré-alongamento.
A produção de força muscular também é dependente da temperatura. Quando ocorre um aumento da temperatura, ocorre um aumento na velocidade de condução no sarcolema, aumento da freqüência de excitação, atividade metabólica e enzimática, e aumento da elasticidade dos componentes elásticos. Essas alterações aumentam a eficiência muscular. Por outro lado à redução da temperatura tem efeito oposto, principalmente decorrente da acidose muscular (redução do PH).
A capacidade de um músculo contrair e relaxar depende da disponibilidade de ATP. Com uma quantidade adequada de oxigênio e nutrientes, poderá haver ATP em quantidade adequada e permitirá tetanias de baixa intensidade por longo tempo.
Entretanto, se a freqüência de tetanias for alta o suficiente para superar a produção de ATP. As respostas de tetania ficarão cada vez mais fracas. Isto é chamado de fadiga. Quando a contração começa, há utilização de ATP providos pela fosfocreatina, enquanto ocorre um rápido aumento da fosforilação oxidativa e da glicólise. Quando o exercício é intenso, o ATP é utilizado rapidamente e a capacidade da célula em substituir o ATP se torna limitada, uma vez que, a atividade enzimática dessa produção é relativamente lenta.
Fibras musculares individuais também exibem diferenças distintas na suas taxas de contração, desenvolvimento de tensão de suscetibilidade à fadiga. Foram identificados três tipos distintos de fibras musculares. As fibras do tipo I são caracterizadas por baixa miosina ATPase, tendo um tempo de contração lento. Apresenta alto conteúdo de mitocôndria com alto potencial para atividade oxidativa. São resistentes à fadiga, uma vez que, ocorre maior fluxo na entrega de oxigênio e nutrientes, suficientes para manter o ritmo de oferta de ATP.
As fibras musculares do tipo II são divididas em dois subgrupos II a e II b. As fibras II a são consideradas intermediárias entre o tipo I e tipo II b. Elas apresentam um tempo rápido de contração combinado com uma capacidade moderada para as atividades aeróbia e anaeróbia.
As fibras II b dependem principalmente da atividade glicolítica (anaeróbia) para produção de ATP. Apresenta poucos capilares e pouca mioglobina. São pouco resistentes à fadiga, mas podem produzir grande tensão.
O tipo de fibra muscular é determinado pelo nervo que a inerva. A característica da inervação por sua vez, depende da função do músculo. O músculo sóleo, por exemplo, tem uma importante função postural, e tem alta porcentagem de fibras do tipo I. Mas, outros músculos são exigidos em diferentes circunstancias e apresentam uma mistura dos três tipos de fibras.
Quando um músculo exerce baixa tensão, as unidades motoras compostas de fibras do tipo I são ativadas. À medida que for necessária maior força, as fibras do tipo II a e eventualmente as fibras do tipo II b são ativadas.
Em atletas de alto nível são encontrados diferentes percentuais nos tipos de fibras. Corredores de corridas rápidas apresentam altos percentuais de fibras tipo II, enquanto corredores de longas distâncias apresentam porcentagem mais alta de fibras do tipo I.
O desuso e a imobilização têm efeitos prejudiciais em fibras musculares. Estes efeitos incluem perda de resistência e força e atrofia muscular. Mudanças bioquímicas acontecem e afetam a produção de energia aeróbia e anaeróbia. O tipo de fibra e o comprimento muscular durante a imobilização. A imobilização em uma posição alongada tem um efeito menos danoso ao músculo. Nessa posição pode ocorrer um aumento do número de sarcômeros em série, enquanto que na posição encurtada ocorre uma diminuição do número de sarcômeros em série. A posição encurtada também favorece maior deposição de tecido conjuntivo.
As fibras do tipo I parecem ser mais susceptíveis à imobilização, possivelmente devido à diminuição do potencial enzimático para a atividade oxidativa. As fibras do tipo I parecem requerer impulsos neurais mais freqüentes, e estes estão reduzidos durante a imobilização.
O treinamento físico com objetivo de aumentar a força muscular é bastante eficiente. Ele aumenta a área de secção transversa das fibras musculares e, portanto, aumenta a capacidade de gerar força. Estímulos elétricos também são capazes de aumentar a força muscular e impedir acúmulo de tecido conjuntivo quando o músculo está imobilizado.
O treinamento físico com o objetivo de aumentar o comprimento muscular por meio dos exercícios de alongamento, também é eficiente. As adaptações a esse treinamento são desde aumento no número de sarcômeros em série e também permitem maior absorção de energia nos elementos contráteis e elásticos.
As bases morfológicas e funcionais aqui abordadas são parte da essência da atividade fisioterapeutica. É por meio do treinamento dos músculos que será possível estabilizar articulações, aumentar a amplitude de movimento, recuperar a capacidade funcional e controle da postura. Portanto, para que o fisioterapeuta exerça seu papel integralmente, é fundamental o conhecimento deste tema.
Os músculos e a gravidade são os principais produtores do movimento humano. Os músculos são usados para manter uma posição, levantar ou abaixar uma parte do corpo, desacelerar movimentos e para gerar velocidade no corpo ou em um objeto. Os músculos também são responsáveis por gerar forças compressivas nas articulações aumentando sua estabilidade. Os músculos são arranjados de forma que possam contribuir individualmente ou coletivamente para produzir um movimento muito pequeno ou um muito amplo e potente. Eles movimentam os ossos nas articulações e dão sustentação à postura do corpo contra forças. Portanto, é um dos principais objetos de trabalho do fisioterapeuta, sendo essencial a compreensão de sua base estrutural e funcional.
O músculo esquelético é o mais abundante no corpo humano com cerca de 40% do peso do corpo. A unidade estrutural do músculo esquelético é a fibra muscular, que consiste de muitas miofibrilas revestidas de uma membrana chamada de sarcolema. O sarcolema é conectado com as linhas Z por proteínas ricas em vinculina e distrofina.
A miofibrila é composta de vários sarcômeros que contêm filamentos de actina, miosina, titina e nebulina. Cada fibra muscular é envolvida por um tecido conjuntivo chamado de endomísio. As fibras musculares são organizadas em feixes, chamados fascículos, que são envolvidos por uma bainha de tecido conjuntivo conhecida como perimísio. O músculo é composto por vários fascículos envolvidos por uma fáscia de tecido conjuntivo fibroso chamado epimísio.
Geralmente, a extremidade de cada músculo é conectada aos ossos por tendões, os quais não possuem propriedade contrátil ativa. Os tendões são contínuos as fibras de colágeno do perimísio e epimísio. Os músculos formam o componente contrátil e os tendões os componentes elásticos em série. Já o endomísio, perimísio, epimísio e o sarcolema compõem os componentes elásticos em paralelo.
Quando o músculo é estirado passivamente, ocorre aumento de tensão e há armazenamento de energia pelos elementos elásticos em série e em paralelo. Por outro lado, quando cessa a tensão a energia é liberada. A possibilidade de distender e a elasticidade dos componentes elásticos são importantes para assegurar a transmissão suave da tensão durante a contração; retorno à posição original após a contração; prevenção de sobre estresse nos elementos contráteis; e a absorção e dissipação de energia.
Cada fibra muscular é composta de muitas miofibrilas acomodadas em paralelo umas as outras no sarcoplasma. Um padrão estriado se repete nas miofibrilas e que é referente aos sarcômeros. O sarcômero é a unidade funcional do sistema contrátil. Ele é composto de filamentos finos (composto de actina); filamentos grossos (composto de miosina); filamentos elásticos (compostos de titina); e componentes inelásticos (compostos de nebulina e titina).
A actina tem a forma de uma hélice dupla (fileira de gotas) na banda I. Duas proteínas adicionais estão junto à actina. A Tropomiosina é uma cadeia longa de polipeptídios que se acomoda nos sulcos entre as hélices da actina. A Troponina é uma molécula globular anexa periodicamente a tropomiosina.
A actina é conectada da linha Z (elemento que une os filamentos finos de sarcômeros adjacentes) até o centro do sarcômero sobrepondo-se aos filamentos grossos.
A miosina (filamento grosso) fica situada na região central do sarcômero, onde seu arranjo paralelo dá origem à banda escura (banda A). Ela é composta de moléculas com cauda e cabeça. Elas estão dispostas em direções diferentes a partir de cada metade do filamento. No centro do filamento há um espaço livre de cabeças (zona H). A titina liga a miosina à linha Z, se estendendo até a linha M (no centro da zona H, onde há união dos filamentos grossos), mantendo a posição central desse filamento. Da linha Z até a miosina a titina tem a aparência de uma mola, útil no desenvolvimento da força muscular passiva no estiramento. Ela também pode servir como um molde para a formação de miosina.
Os filamentos de nebulina se estendem da linha Z até os filamentos de actina, e podem servir como um molde para a formação de actina.
Junto às miofibrilas encontra-se acomodada uma rede organizada de túbulos e bolsas conhecida como retículo sarcoplasmático. Essa rede tende a alargar-se e fundir-se no nível das bandas A e I, formando as cisternas terminais e o sistema T (sistema transverso tubular).
A teoria proposta por Huxley parte do pressuposto que o sarcômero encurta por meio do deslizamento dos filamentos grossos e finos mantendo seu comprimento original. O deslizamento ocorreria por meio de pontes cruzadas (ligações entre actina e miosina) com posterior movimento angular da cabeça de miosina (similar ao remo), que depois se separa e se junta a outro local para novo movimento angular da cabeça.
As ligações cruzadas são assincrônicas, com cerca de 50% das ligações gerando força, enquanto os outros 50% manterão a posição quando os primeiros 50% se separarem.
O Cálcio é o mecanismo chave para a contração muscular. Se disponível, ocorre contração, mas se ausente a contração cessa. A presença do cálcio é regulada por eventos elétricos no sarcolema. O neurônio motor estimula o músculo (ao nível da junção neuromuscular) e o potencial de ação propaga-se ao longo de todo o sistema T. Um neurônio motor e as fibras musculares inervadas por ele formam a unidade motora, e respondem a lei do tudo ou nada (despolariza ou não despolariza). A quantidade de fibras para cada neurônio em um músculo depende de sua função, quanto maior a necessidade de precisão da ação muscular menor a quantidade de fibras musculares para cada neurônio. Os terminais axônicos acomodam-se nas fendas do sarcolema. Esses terminais estão repletos de vesículas sinápticas, as quais liberam a acetilcolina para transmitir o potencial de ação gerado no nervo para o sarcolema, e por fim, a liberação do cálcio.
A resposta mecânica ao estímulo nervoso é chamada de tetania. Existe um período entre o estímulo e a tetania, chamado de latência. Essa latência ocorre devido ao tempo necessário para eliminar a “folga” dos elementos elásticos. Ele também é chamado de atraso neuromecânico (também chamado de eletromecânico, no inglês electromechanical delay).
Do início da tetania até o seu pico temos o tempo de contração, e do pico até o final da tetania temos o tempo de relaxamento.
O acréscimo de resposta mecânica a estímulos sucessivos é chamado de somação. Se houve um segundo estímulo antes do término da tetania, haverá nova tensão, e possivelmente maior do que a primeira. Quando o segundo estímulo é logo após o primeiro (pequeno intervalo), há aumento da tensão resultante é maior do que seria com apenas um estímulo. Caso haja uma seqüência de estímulos com pequeno intervalo, ou seja, a freqüência é alta, haverá uma contração tetânica máxima, mantendo a tensão de pico.
As diferenças na gradação, freqüência de estímulo e número de unidades ativadas produzem os movimentos suaves que costumamos observar. A contração muscular exerce uma força sobre os ossos conhecida como tensão muscular. A força externa aplicada no músculo é chamada de resistência ou carga. A tensão muscular gera um efeito de rotação nas articulações chamado de momento ou torque. Ele é calculado como o produto da força muscular e a distância perpendicular da aplicação da força e o centro do movimento.
Contrações musculares e o trabalho muscular podem ser classificados de acordo com a tensão muscular e a resistência a ser superada. O trabalho estático ocorre quando nenhum trabalho mecânico é executado, mas há trabalho muscular (gasto de energia) por meio de uma contração isométrica.
O trabalho dinâmico ocorre quando um trabalho mecânico é produzido, e o movimento articular é gerado por uma das formas de contração muscular. A contração concêntrica ocorre quando os músculos geram tensão suficiente para superar a resistência do segmento do corpo e os músculos se encurtam. A contração excêntrica ocorre quando os músculos não geram tensão suficiente e são superados pela carga externa, de modo que eles se alongam (afastam origem e inserção). A contração isocinética é uma forma de trabalho muscular onde a velocidade de alongamento ou encurtamento se mantém constantes. Este tipo de contração é obtido por meio de equipamentos computadorizados. A contração isotônica é usada para definir a contração muscular com a tensão constante ao longo da amplitude articular. O uso desse termo não leva em conta que durante a amplitude de movimento ocorre mudanças no braço da alavanca, modificando também a tensão. Portanto, uma contração isotônica no verdadeiro sentido não existe, mas o termo muitas vezes é usado para definir uma contração que provoca movimento articular.
A tensão varia com o tipo de contração. As contrações isométricas produzem maior tensão do que contrações concêntricas. Contrações excêntricas podem gerar maior tensão do que contrações isométricas. Isto é aceito levando em conta a tensão suplementar produzida pelos elementos elásticos.
A classificação funcional da atividade muscular é necessária para melhor compreender como os músculos estão atuando. Agonista é o termo usado para denominar o músculo ou grupo muscular responsável por determinado movimento. O grupo muscular ou músculo responsável por realizar o movimento oposto ao agonista é chamado de antagonista. Quando um músculo auxilia o agonista na execução de um movimento é chamado de sinergista. Os fixadores são músculos que atuam com certo grau de sinergismo, uma vez que eles são músculos que produzem estabilidade em uma articulação de forma a favorecer o movimento em outra articulação.
Dependendo da posição e direção da resistência a superar ocorre diferenças na atividade do músculo, se concêntrica, excêntrica ou ainda não participa da atividade.
A produção de força ou tensão de um músculo depende do comprimento em que o mesmo está quando estimulado. A tensão máxima é produzida quando o músculo está próximo do seu comprimento de repouso. Quando o músculo está em uma posição encurtada, a tensão cai lentamente a princípio e depois mais rapidamente. Quando o músculo está em uma posição de alongamento, a tensão progressivamente diminui. Isto está relacionado com as alterações estruturais no sarcômero. Na posição de repouso, os filamentos de actina e miosina estão sobrepostos ao longo de todo o seu comprimento e o número de pontes cruzadas é máximo. Na posição alongada, haverá menos haverá menos pontes cruzadas e a tensão será menor. Na posição encurtada os filamentos estão sobrepostos nos terminais opostos do sarcômero, diminuindo a tensão.
A velocidade de encurtamento é maior quando a carga externa zero, mas à medida que a carga aumenta, a velocidade diminui. Quando a carga é aumentada ainda mais, o músculo alonga-se e contrai excentricamente. Com o aumento da carga ocorre maior velocidade de alongamento. Portanto, a relação carga-velocidade da contração excêntrica é inversa à da contração concêntrica.
A força ou tensão é proporcional ao tempo de contração. A força é maior se há tempo necessário para a tensão produzida ser transmitida para os elementos elásticos. O músculo que apresenta fibras musculares com muitos sarcômeros em série, terá maior velocidade e excursão (alcance de trabalho). O músculo que apresenta maior número de sarcômeros em paralelo terá maior produção de força.
O músculo realiza maior trabalho se for alongado e logo em seguida realizar uma contração concêntrica, a partir de uma contração isométrica. Os exercícios pliométricos são um bom exemplo desse efeito do pré-alongamento.
A produção de força muscular também é dependente da temperatura. Quando ocorre um aumento da temperatura, ocorre um aumento na velocidade de condução no sarcolema, aumento da freqüência de excitação, atividade metabólica e enzimática, e aumento da elasticidade dos componentes elásticos. Essas alterações aumentam a eficiência muscular. Por outro lado à redução da temperatura tem efeito oposto, principalmente decorrente da acidose muscular (redução do PH).
A capacidade de um músculo contrair e relaxar depende da disponibilidade de ATP. Com uma quantidade adequada de oxigênio e nutrientes, poderá haver ATP em quantidade adequada e permitirá tetanias de baixa intensidade por longo tempo.
Entretanto, se a freqüência de tetanias for alta o suficiente para superar a produção de ATP. As respostas de tetania ficarão cada vez mais fracas. Isto é chamado de fadiga. Quando a contração começa, há utilização de ATP providos pela fosfocreatina, enquanto ocorre um rápido aumento da fosforilação oxidativa e da glicólise. Quando o exercício é intenso, o ATP é utilizado rapidamente e a capacidade da célula em substituir o ATP se torna limitada, uma vez que, a atividade enzimática dessa produção é relativamente lenta.
Fibras musculares individuais também exibem diferenças distintas na suas taxas de contração, desenvolvimento de tensão de suscetibilidade à fadiga. Foram identificados três tipos distintos de fibras musculares. As fibras do tipo I são caracterizadas por baixa miosina ATPase, tendo um tempo de contração lento. Apresenta alto conteúdo de mitocôndria com alto potencial para atividade oxidativa. São resistentes à fadiga, uma vez que, ocorre maior fluxo na entrega de oxigênio e nutrientes, suficientes para manter o ritmo de oferta de ATP.
As fibras musculares do tipo II são divididas em dois subgrupos II a e II b. As fibras II a são consideradas intermediárias entre o tipo I e tipo II b. Elas apresentam um tempo rápido de contração combinado com uma capacidade moderada para as atividades aeróbia e anaeróbia.
As fibras II b dependem principalmente da atividade glicolítica (anaeróbia) para produção de ATP. Apresenta poucos capilares e pouca mioglobina. São pouco resistentes à fadiga, mas podem produzir grande tensão.
O tipo de fibra muscular é determinado pelo nervo que a inerva. A característica da inervação por sua vez, depende da função do músculo. O músculo sóleo, por exemplo, tem uma importante função postural, e tem alta porcentagem de fibras do tipo I. Mas, outros músculos são exigidos em diferentes circunstancias e apresentam uma mistura dos três tipos de fibras.
Quando um músculo exerce baixa tensão, as unidades motoras compostas de fibras do tipo I são ativadas. À medida que for necessária maior força, as fibras do tipo II a e eventualmente as fibras do tipo II b são ativadas.
Em atletas de alto nível são encontrados diferentes percentuais nos tipos de fibras. Corredores de corridas rápidas apresentam altos percentuais de fibras tipo II, enquanto corredores de longas distâncias apresentam porcentagem mais alta de fibras do tipo I.
O desuso e a imobilização têm efeitos prejudiciais em fibras musculares. Estes efeitos incluem perda de resistência e força e atrofia muscular. Mudanças bioquímicas acontecem e afetam a produção de energia aeróbia e anaeróbia. O tipo de fibra e o comprimento muscular durante a imobilização. A imobilização em uma posição alongada tem um efeito menos danoso ao músculo. Nessa posição pode ocorrer um aumento do número de sarcômeros em série, enquanto que na posição encurtada ocorre uma diminuição do número de sarcômeros em série. A posição encurtada também favorece maior deposição de tecido conjuntivo.
As fibras do tipo I parecem ser mais susceptíveis à imobilização, possivelmente devido à diminuição do potencial enzimático para a atividade oxidativa. As fibras do tipo I parecem requerer impulsos neurais mais freqüentes, e estes estão reduzidos durante a imobilização.
O treinamento físico com objetivo de aumentar a força muscular é bastante eficiente. Ele aumenta a área de secção transversa das fibras musculares e, portanto, aumenta a capacidade de gerar força. Estímulos elétricos também são capazes de aumentar a força muscular e impedir acúmulo de tecido conjuntivo quando o músculo está imobilizado.
O treinamento físico com o objetivo de aumentar o comprimento muscular por meio dos exercícios de alongamento, também é eficiente. As adaptações a esse treinamento são desde aumento no número de sarcômeros em série e também permitem maior absorção de energia nos elementos contráteis e elásticos.
As bases morfológicas e funcionais aqui abordadas são parte da essência da atividade fisioterapeutica. É por meio do treinamento dos músculos que será possível estabilizar articulações, aumentar a amplitude de movimento, recuperar a capacidade funcional e controle da postura. Portanto, para que o fisioterapeuta exerça seu papel integralmente, é fundamental o conhecimento deste tema.
Referências
Bibliográficas
1- Nordin, M; Frankel, VH. Biomecânica básica do sistema musculoesquelético. 3.ed. Rio de Janeiro: Guanabara Koogan, 2003.
1- Nordin, M; Frankel, VH. Biomecânica básica do sistema musculoesquelético. 3.ed. Rio de Janeiro: Guanabara Koogan, 2003.
2-
Smith, LK; Weiss, EL; Lehmkuhl, LD. Cinesiologia Clínica de
Brunnstrom. 5 ed. São Paulo: Manole, 1997.
3-
Hamill, J.; Knutzen, K.M. Bases biomecânicas do movimento humano. 2
ed. Editora Manole, SP, 2008.
4-
Marques, AP. Cadeias Musculares – Um Programa para Ensinar
Avaliação Fisioterapêutica Global. São Paulo: Manole, 2000.










